


Cell Metabolism:多能干细胞命运中的营养素
2021年10月,加利福尼亚大学洛杉矶分校等单位的相关研究人员在《Cell Metabolism》(IF: 27.3)上发表了题为“Nutrients in the fate of pluripotent stem cells”的综述文章,评估了影响多能干细胞命运的营养素及其代谢转化的可用数据,还讨论了在这一迅速扩大的、日益具有基础性和实践重要性的领域中有待研究的关键问题。

在考虑多能干细胞(PSCs)的命运时,我们将多能/自我更新干细胞定义为表达核心多能性转录因子(包括OCT4、NANOG、SOX2等)的细胞,并将分化定义为核心多能性转录因子的沉默以及谱系特异性生物标记物的获得。研究表明,无论是否有细胞内加工,供应的营养素在PSC特性和细胞命运转换中都具有积极的指导作用。例如,通过糖酵解、三羧酸(TCA)循环和单碳代谢产生的营养物质代谢通量产生中间代谢物,用于修饰PSC表观基因组、染色质结构和基因表达的反应。
除了将PSC产生的中间代谢物与表观基因组修饰联系起来的研究外,关于营养素(包括糖、氨基酸、脂质等)在控制PSC命运中的作用,还有许多有趣的问题。例如,最近的结果表明,营养素也可以通过合成代谢或分解代谢途径的代谢通量以外的机制影响PSC的命运。对这些研究的一种潜在解释是在控制 PSC 命运的触发器之前或同时,特定的营养素在微环境中发出信号条件。
目前的研究表明,相同的营养素可以启动或支持 PSC 的命运,其不同的影响取决于环境,例如多能性状态、分化线索、培养基条件和定时添加或剥夺营养素(表 1)。
表1. 引发不同 PSC 命运的营养素示例

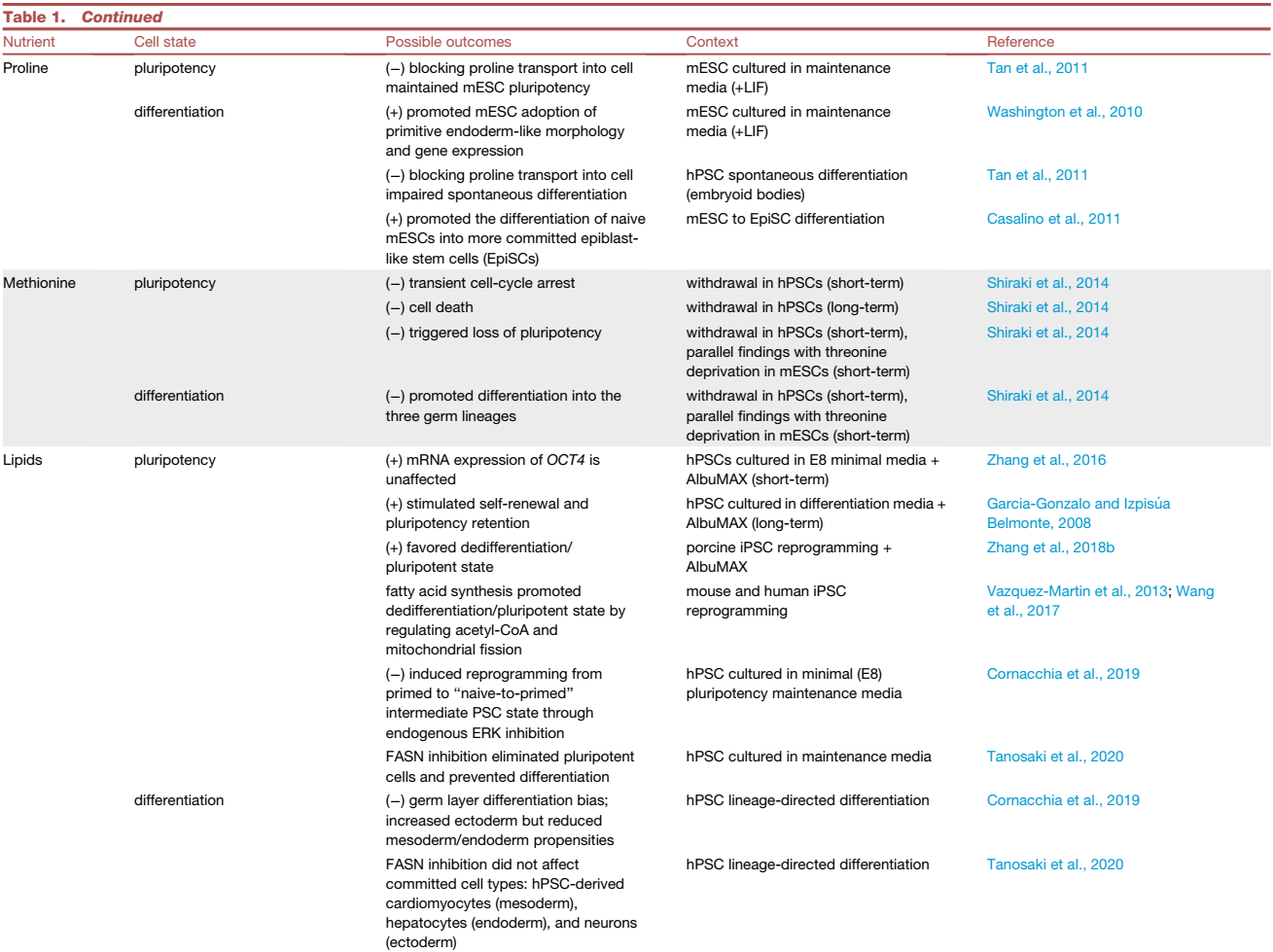
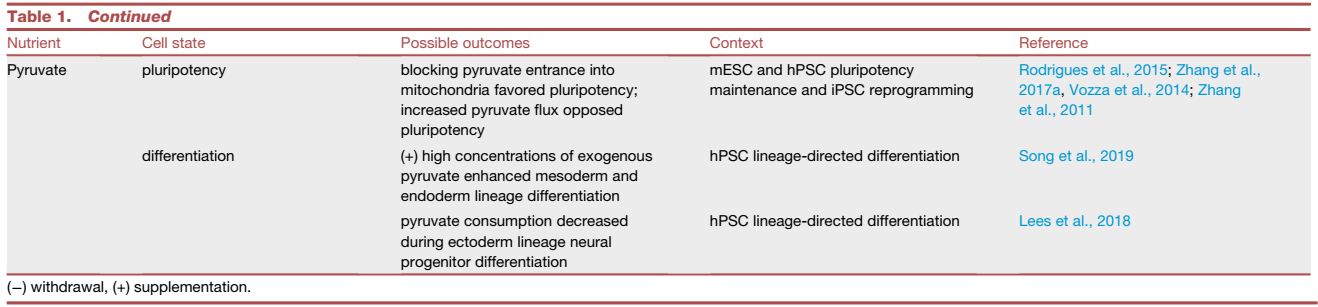
特定的营养素缺乏或添加不会在体内自然发生,但体外营养素变化提供了一个有价值的实验工具。例如,从PSC培养基中定时提取谷氨酰胺显示出对PSC命运的不同影响。短期停用谷氨酰胺消除了TCA循环回补中高度依赖谷氨酰胺的细胞,使存活的小鼠胚胎干细胞(mESCs)和人类PSCs(hPSCs)富含多能转录因子OCT4和SOX2的表达。相反,也有研究表明,高水平的谷氨酰胺代谢阻止了OCT4的降解,并保持了hPSC的自我更新。长期缺乏谷氨酰胺也会导致更多的氧化性PSC因TCA循环活性降低和氧化磷酸化(OXPHOS)而死。在这种情况下,谷氨酰胺停药提供了一种消除PSC并富集hPSC衍生心肌细胞的策略或增强内皮细胞的血管生成能力。综合结果表明,在补充谷氨酰胺后的短期停用可以消除更多的氧化性PSC,并产生更同质的具有更高多能性转录因子表达的PSC。因此,在PSC培养中控制谷氨酰胺暴露的时间可以控制PSC或非多能细胞是否富集。
葡萄糖是哺乳动物细胞的另一种关键营养素,似乎也控制着 PSC 结果。在小鼠早期胚胎发生中,在致密的 8 细胞桑椹胚阶段,虽然葡萄糖是必需的,但糖酵解是可有可无的。在桑椹转化为滋养外胚层 (TE) 和囊胚内细胞团(ICM)的过程中,这是第一个命运规范,葡萄糖依赖性信号传导指导胚外 TE 的形成,而不是ICM。 这些离体小鼠研究显示葡萄糖输入支持戊糖磷酸途径 (PPP) 核苷酸生产和己糖胺生物合成途径 (HBP) 信号,激活 TE 特异性转录因子 CDX2。 在葡萄糖缺乏培养基中培养显示 TE 生物标志物的表达降低,而 ICM 特异性 OCT4 和 NANOG 表达未受影响。因此,最早的细胞命运规范显示了发育线索,至少部分是由营养可用性和营养敏感信号控制的。
研究表明,额外的营养素,如抗坏血酸、脯氨酸和甲硫氨酸,也能调节多能性。在人类 iPSC 集落中,抗坏血酸通过增强组蛋白去甲基化酶 JARID1 表达积极促进多能性并抑制自发分化。抗坏血酸在干细胞分化中有这独特而多样的作用。例如,在心脏分化提示下,抗坏血酸通过增加胶原合成增加心脏祖细胞的增殖来促进心脏分化,但仅在分化早期补充到培养基中。
添加脯氨酸促进 mESC 采用原始外胚层样形态,并提高与原始外胚层相关基因的表达。脯氨酸通过氨基酸转运蛋白 SNAT2 导入。用竞争性底物抑制 SNAT2 会阻止脯氨酸转运,维持 mESC 多能性,并损害自发性胚状体 (EB) 三系分化。脯氨酸补充还促进幼稚的 mESC 分化为更成熟的外胚层样干细胞 (EpiSCs)。与脯氨酸一样,甲硫氨酸在多能性和分化中发挥作用。在 hPSC 中,S-腺苷甲硫氨酸(SAM)支持的组蛋白甲基化依赖于甲硫氨酸而不是苏氨酸代谢。hPSC 中的短期甲硫氨酸剥夺会降低细胞内 S-甲基-50-硫腺苷 (MTA)(一种甲硫氨酸前体)和 SAM,后者由甲硫氨酸生成(图 1)。这种短期操作会导致短暂的细胞周期停滞,而长期缺乏甲硫氨酸会导致细胞死亡。在长期甲硫氨酸剥夺期间补充 MTA 或 SAM 可挽救 hPSC 增殖,表明甲硫氨酸衍生的代谢物对于持续的 hPSC 自我更新至关重要。短期甲硫氨酸剥夺也可引发多能性丧失和 hPSC 分化为三个胚系。脯氨酸和甲硫氨酸如何影响不同发育阶段的多能性状态将是未来研究的一个有趣主题。
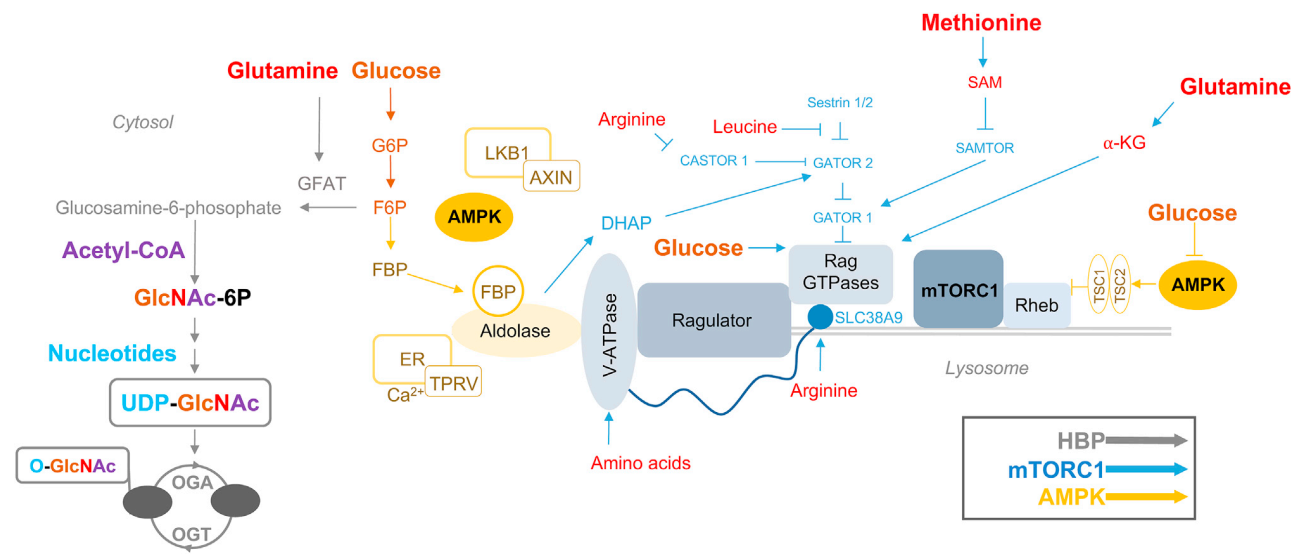
图1. 营养感应通路影响多能干细胞的命运
总之,这些研究表明,某些营养素(如谷氨酰胺、葡萄糖、抗坏血酸、脯氨酸和甲硫氨酸)的可用性会导致不同的、有时是相反的 PSC 命运结果(表 1)。 一个悬而未决的问题是某些营养素是否提供足够的指导信号来控制特定的发育阶段或转变。例如,特定的营养素能否替代某些信号细胞因子、生长因子、激素或重编程因子以实现或维持多能性,或促进谱系特异性分化?
特定酶活性和转运蛋白对营养物质的受控氧化和细胞内路由可以诱导或维持多能性,表明营养物质处理途径配置调节 PSC的命运。脂质代谢,例如脂肪酸 (FA) 氧化,对于早期小鼠和人类胚胎发育至关重要。事实上,脂质信号传导与多能性有关,将脂质可用性与 PSC 命运相关联的发现表明,脂质的缺失会通过 FA 生物合成触发葡萄糖路由升高,从而支持多能性。
细胞内丙酮酸路径可以发生在 (1) 通过乳酸脱氢酶 (LDH) 产生乳酸,产生 NAD+ 以维持糖酵解通量,(2) 通过丙酮酸脱氢酶 (PDH) 产生乙酰辅酶 A 用于 TCA 循环氧化,(3) 通过丙酮酸产生草酰乙酸 羧化酶 (PCB) 来补充 TCA 循环中间体,以及 (4) 定位于细胞核以调节表观基因组(图 1)。PDH 抑制阻止丙酮酸进入 TCA 循环,支持小鼠 iPSC 重编程和活力,而进入线粒体的丙酮酸流量增加会降低多能性生物标志物表达,减少 PSC 自我更新,并降低 PSC 活力。分化后,PDH 基因表达迅速下降,但 PDH 蛋白水平保持不变。 相比之下,ATP-柠檬酸裂解酶 (ACYL) 和乙酰辅酶 A 合成酶 (ACCS2) 为支持多能性的组蛋白乙酰化反应提供丙酮酸衍生的乙酰辅酶 A,在分化时迅速减少。这种代谢酶表达的命运依赖性模式表明,在多能性退出后,丙酮酸路线的层次转向线粒体 TCA 循环氧化以支持 PSC 分化。此外,多项研究表明,代谢酶的激活和转运蛋白的表达先于基因表达和 PSC 身份的变化,这可能是为细胞身份转换做准备的一种机制。
多能性中的mTOR
表2. 营养敏感信号转导通路

多能性中的AMPK
多能性中的糖基化
O-GlcNAcylation 在调节 PSC 命运方面存在另一层复杂性,因为营养水平与 O-GlcNAcylation 之间可能不存在线性关系。多项研究报告称,葡萄糖、谷氨酰胺和氨基酸没有显示与蛋白质 O-GlcNAc 水平的剂量依赖关系。这表明 O-GlcNAcylation 的营养调节可能与 PSC 环境有关,这是营养控制 PSC 命运的反复出现的主题。例如,特定营养物质的可用性是增加还是减少多能性转录因子的 O-GlcNAcylation 水平可能取决于分化线索或 PSC 状态,而不是整体 HBP 通量。
过去二十年的研究使大多数研究人员相信代谢在控制 PSC 命运中的积极作用,但仍有许多问题。这些包括营养物质在 PSC 命运中的作用,在细胞身份转变之前或期间。例如,特定的养分可用性是启动还是仅加强 PSC 命运“决定”?外源性营养素与细胞内源性营养素需求在指导或加强 PSC 命运方面有何作用?技术进步以及单细胞代谢组学质谱检测和灵敏度的进步,可以揭示体内和体外 PSC 身份转换过程中不同生态位细胞类型消耗、产生和分泌的营养物质。
未来研究的一个实际领域是确定触发影响 PSC 命运的营养信号的阈值浓度。这些研究的结果可能对孕产妇饮食和/或糖尿病如何影响发育产生影响。例如,由于氧化和内质网应激,过高浓度的葡萄糖不利于 PSC 衍生的神经谱系生成,但也会促进 PSC 衍生的心肌细胞生成。 此外,高浓度的丙酮酸通过 AMPK 激活的信号传导促进中胚层而非外胚层分化。


