Nature:抑制脂肪酸氧化可使成年小鼠心脏再生
2023年9月,德国马克斯·普朗克心肺研究所等单位的相关研究人员在《Nature》(IF: 64.8)上发表了题为“Inhibition of fatty acid oxidation enables heart regeneration in adult mice”的研究论文,发现通过重塑心肌细胞(CMs)成熟状态的代谢方式,可逆转其表观遗传性状,解除成熟心肌细胞的增殖障碍,从而实现心脏再生。

-
心肌细胞中脂肪酸氧化的阻断可以提高对缺氧的抵抗力,刺激心肌细胞增殖。
-
肉毒碱棕榈酰转移酶Cpt1b突变心肌细胞中的α-酮戊二酸(αKG)积累可激活组蛋白赖氨酸去甲基酶KDM5。
-
活化的KDM5降低 H3K4me3水平并将心肌细胞转化到不太成熟的状态,从而促进增殖。
研究背景:
在出生后,CMs的能量代谢就从糖酵解转变为氧化代谢,在这个过程中,线粒体稳态、细胞结构和电生理特性都将发生重塑。与此同时,大多数CMs退出细胞周期,发生肥大性生长以及细胞通讯方式的变化。由于高代谢活性,CMs的出生后成熟和氧化DNA损伤形成了一种天然的屏障,阻止CM细胞分裂。CMs的代谢和结构成熟是紧密交织在一起的。这两个过程中的任何一个的逆转都可能在 CMs中重新建立增殖能力。然而,将代谢重塑与转录和结构改变联系起来,限制成熟CMs增殖的机制尚不清楚。
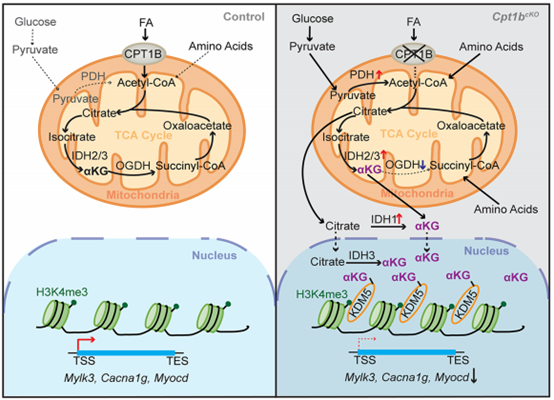
不同代谢物(作为各种染色质修饰物的必需辅因子或底物)浓度的变化,可能会将代谢过程与染色质状态和基因活性联系起来。αKG是一种三羧酸循环的中间产物,是几种双加氧酶活性所必需的。αKG依赖性KDMs通过调节H3K4me3和H3K9me3组蛋白修饰水平,在心脏发育、CMs成熟和心脏功能中发挥关键作用。目前关于αKG在染色质重构中的作用的信息很少,并且αKG在促进心脏再生中的潜在作用至今尚未被研究。
RNA测序(RNA-seq)数据集的分析显示,在出生后第一周CM成熟过程中,糖酵解和细胞周期进展的几个关键基因的表达水平降低,而与脂肪酸氧化(FAO)和三羧酸循环相关的基因被上调。上调的FAO相关基因包括肉毒碱棕榈酰转移酶Cpt1b的肌肉特异性亚型,但不包括普遍表达的Cpt1a亚型,而这是线粒体摄取脂肪酸和随后的FAO所必需的。
为了确定FAO在体内调节CM增殖和生长的作用,研究人员构建了Cpt1b敲除(Cpt1b cKO)小鼠,其中Cpt1b基因在胚胎阶段的CM中特异性失活。观察发现,与对照小鼠相比,Cpt1b cKO小鼠的体重(BW)、心脏重量(HW)、HW/BW比值和心脏形态未发生变化。然而,由于心肌同心生长,10周龄Cpt1b cKO小鼠的心脏大小、HW和HW/BW增加。值得注意的是, Cpt1b cKO小鼠心脏中CMs的绝对数量增加一倍。此外,Cpt1b cKO小鼠的CM表面积相对适度增加,这表明心肌同时发生增生性和肥厚性生长。形态学评价显示, CM群体显著增加,表明Cpt1b失活后会形成新的CMs,中等大小CMs的比例下降,而较大CMs的数量显著增加,反映了CM表面积的总体增加。这些数据表明,Cpt1b的缺失会刺激CM的增殖。
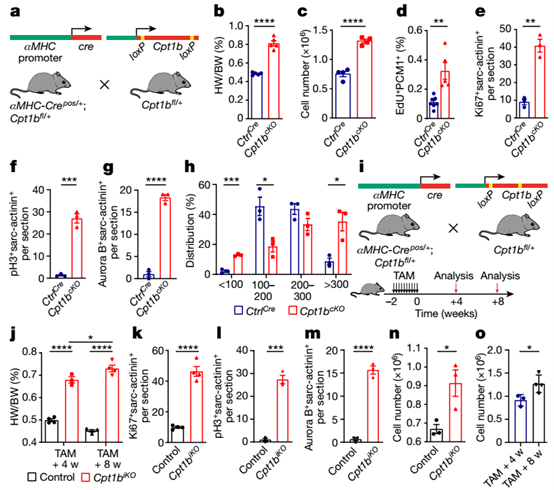
Cpt1b失活诱导CMs的增生性和肥大性生长
为了研究Cpt1b失活和随后的CMs增殖是否能够实现心脏再生,研究人员对Cpt1b敲除小鼠进行了缺血-再灌注(I–R)损伤,该模型与接受支架重建阻塞冠状动脉的人类患者的情况非常相似。结果发现,阻断FAO获得的增殖潜力允许先前存在的CMs重新进入细胞周期并有助于心脏再生。组织学分析显示,与对照动物相比,Cpt1b敲除小鼠的纤维瘢痕面积显著减少,在I–R手术后4周,心脏功能显著恢复,几乎达到了损伤前的水平,证明Cpt1b失活促进心脏再生。

Cpt1b介导的FAO废除可防止I–R损伤,并使心脏再生
接下来,研究人员探究了FAO的阻断如何重新编程CMs的代谢。代谢通量和Seahorse分析表明,长链脂肪酸的利用在Cpt1bcKO CMs中被有效抑制。此外,靶向代谢组分析显示,来源于中链和长链脂肪酸的酰基肉碱水平在Cpt1b缺陷型CMs中显著降低,而游离肉碱的细胞内水平显著升高。而细胞内乙酰辅酶A和三羧酸循环的大多数代谢产物,如柠檬酸、琥珀酸和苹果酸的水平没有显著改变,表明在没有FAO的情况下,CMs使用不同的形式产生能量。通过对氨基酸代谢产物的分析及代谢流实验验证,Cpt1b缺陷型CMs中葡萄糖氧化和支链氨基酸(BCAA)分解代谢的增强,有效地补偿了Cpt1b缺陷CMs中脂肪酸衍生的乙酰辅酶A进入三羧酸循环的代谢通量损失。值得注意的是,Cpt1b失活后CMs中的αKG显著增加了近20倍。谷氨酰胺和谷氨酸直接转化为αKG似乎不太可能解释αKG的积累,因为谷氨酰胺和谷氨酸的浓度没有下降。此外,异柠檬酸水平显著降低,同时IDH1、IDH2和IDH3A水平增加,它们催化异柠檬酸转化为αKG。表明FAO阻断后,合成增加和代谢减少协同作用,在CMs中积累αKG。

脂肪酸的线粒体输入受阻重新连接CMs的代谢并提高αKG水平
αKG是组蛋白去甲基化酶的重要辅因子。因此,研究人员接着探究了Cpt1b缺陷CMs中αKG的积累是否影响组蛋白甲基化。使用通过荧光激活细胞分选(FACS)分选的细胞核对不同组蛋白赖氨酸甲基化修饰的分析显示,只有H3K4me3的水平降低。而在Cpt1b失活后,异染色质标记物如H3K9me3、H3K27me3和H4K20me2的水平显著增加,这可能是由于与CMs更成熟状态的转变带来的次级效应。
使用FACS分选的CM核对H3K4me3进行染色质免疫沉淀和测序(Chip-seq)分析,结合ChIP–qPCR验证,证实在与 CM 成熟相关的基因的启动子和基因体中,H3K4me3富集广泛减少,而启动子处的 H3K9me3和 H3K27me3水平保持不变。因此,αKG 的积累激活 H3K4me3特异性去甲基化酶,其随后在心脏分化和成熟所需的基因内消除 H3K4me3,从而将 Cpt1b 缺陷型 CM 转化为更不成熟的、具有增殖能力的状态。
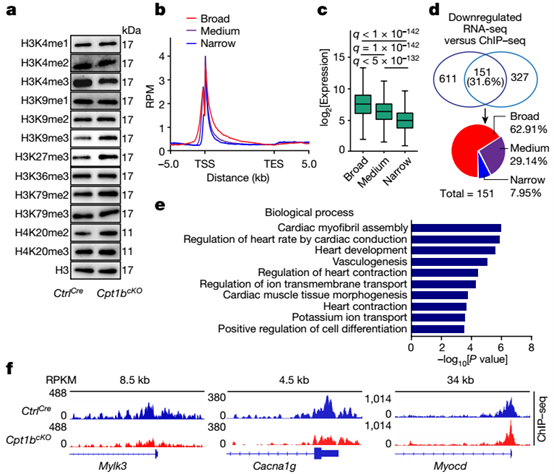
增加的αKG水平诱导H3K4me3去甲基化并降低CM成熟基因的表达
为了证实αKG的积累是阻止CM成熟和促进增殖的关键信号,研究人员用细胞可渗透的αKG处理P0-1 CM。观察到主要在胎儿和新生儿阶段表达的基因水平显著升高,而与CM成熟和分化相关的基因水平降低。用αKG处理也降低了H3K4me3,但不影响H3K9和H4K20甲基化。KDM5抑制剂 CPI-455对 αKG 依赖性作用的药理学抑制表明,αKG 对染色质和基因表达的影响是由 Kdm5基因家族成员传递的。Kdm5b 的过表达证实了这一假设,当与 αKG 处理结合时,Kdm5b 强烈降低 H3K4me3水平并显著增加 CM 细胞周期活性。
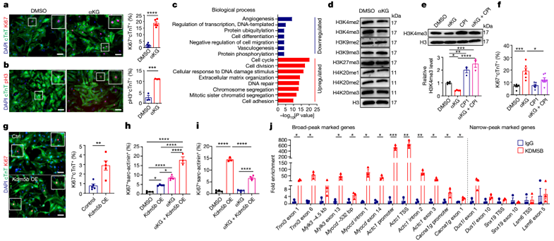
αKG的积累刺激KDM5活性,减弱CMs的成熟并增强其增殖
总的来说,该研究发现单一代谢产物αKG将代谢过程与KDM5介导的转录变化联系起来,这为操纵CMs的命运和功能以及刺激心脏再生提供了许多机会。